热 线:400-0379-440
手 机:15225555522
邮 箱:xiaguangyoule@126.com
邮 编:471000
地 址:河南省洛阳市龙城区春都路53号
新靶标展望之粘附性GPCR海上靶标
粘附性GPCR(Adhesion G protein-coupled receptors,aGPCR)属于GPCR超家族中的一类,在结构上它拥有独特的特征,包括复杂的多结构域的N端以及虽然GPCR一般具有比较明确的药理学性质,但是目前能够靶向aGPCR的小分子还没有进入上市或临床实验阶段。因此,对aGPCR靶点重要性以及开发aGPCR调控分子的方法的汇总对于进一步挖掘该类家族的治疗潜力是很有价值的。
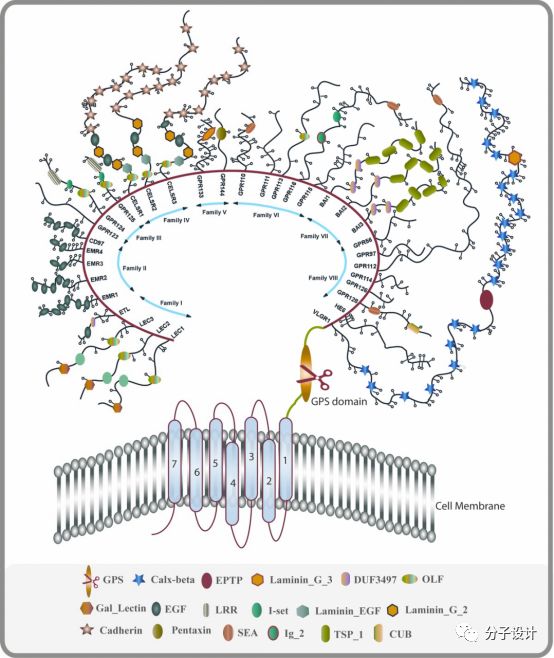
GPCR总共包括800个受体,按照序列和药理学性质可以分为6个受体。aGPCR一开始被认为是B类GPCR中的分泌素家族的一类,但是从结构上看,他有一些独特的特征,包括自水解位点以及长长的细胞外N端部分,它能和除了激素以外的分子结合,因此被单独归为一类。aGPCR的N端通常和参与细胞粘附以及细胞-细胞相互作用的蛋白具有高度的同源度。而其自水解位点可以结合一个名为Stachel的配体分子来激活aGPCR的信号转导。
aGPCR总共33个成员,可以根据跨膜区和胞外区的进化关系可以分为9个类别,海上靶标其命名规则是以ADGR后的字母代表不同的N端结构域。ADGRL是指原来的蛛毒素受体,ADGRE是指包含EGF-TM7受体的表皮因子生长受体结构域,ADGRC是指包含CELSR的钙黏蛋白结构域,ADGRB之前被命名为BAI,包含血栓黏合素结构域,ADGRV之前被命名为VLGR1。其余的aGPCR类也都是按照进化起源和结构域进行命名。
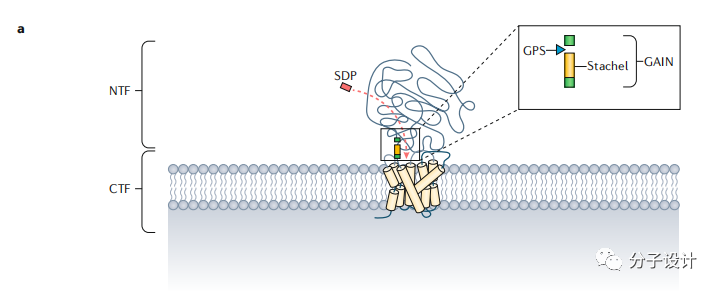
aGPCR是一类十分古老的蛋白,有些成员的直系同源出现在海葵等低等生物中。而包含GPS的GAIN结构域在多个具有不同结构和功能的蛋白中被发现,预示其祖先结构域在进化上很早就已出现。GPS位于跨膜第一个螺旋之前,在自催化的水解作用后,N端片段(NTF),跨膜区和C端片段(CTF)都非共价地偶联在一起。自催化分裂GPCR后,可以结合Stachel,然后激动aGPCR,并将信号传导至G蛋白及下游分子。值得一提是,ADGRF2, ADGRF4和ADGRC1虽然具有GAIN结构域,但不会发生自催化的水解现象,而ADGRA1甚至缺少GAIN和GPS结构域。虽然自催化过程和某些成员(ADGRE5,ADGRG1)的生理活性是相关的,但是ADGRL的信号传感是不需要这个机制的。
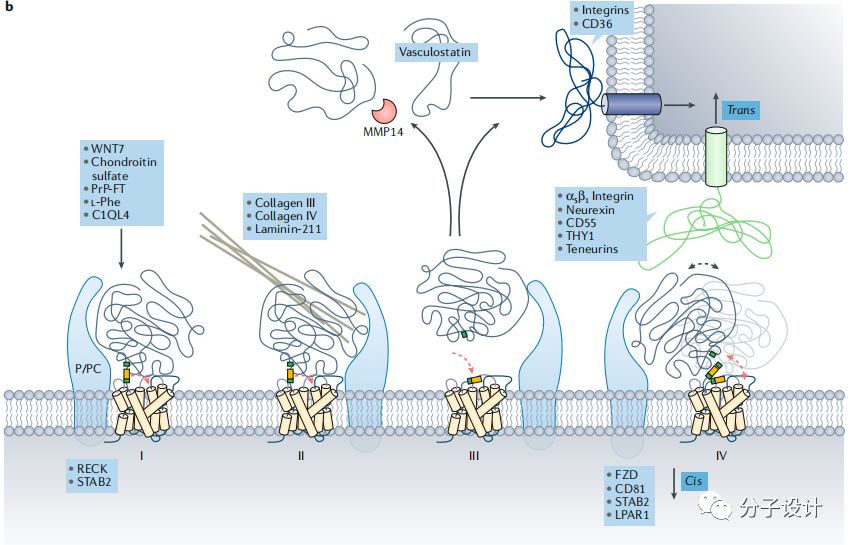
N端的独特不仅仅是因为它们的长度,更是因为它们具有多样的结构域,包括EGF,亮氨酸富集的重复区域,激素受体结构域,免疫球蛋白结构域。这些区域同样存在于其他蛋白中,因此这些受体参与了细胞-细胞和细胞-基质互作。一些已经被确认的和NTF相互作用的分子也证明了这一点。比如,ADGRG1结合小胶质细胞分泌的谷氨酰胺转移酶2,对于细胞黏连和稳定基质就很重要。并且,在细胞外基质(ECM)由谷氨酰胺转移酶2介导的ADGRG1和层黏连蛋白-111使得ADGRG1可以整合小胶质细胞和ECM的信号以此来促进髓鞘的发育和组织修复。同样地,在凋亡细胞表面,ADGRG1对磷脂酰胺的识别可以促进这些细胞的吞噬。海上靶标另一方面,尽管ADGRL的激素结构域和促肾上腺皮质激素释放因子受体1(CRF1)具有结构相似性,但是在激素结合前ADGRL须在结合位点经历构象变化,这也预示着一种新的活化机制。
靶点识别和验证是药物发现的第一步,这对于一个项目最终是否能进入临床是至关重要的,这需要整合各种来源的信息,包括人类,小鼠遗基因组证据和体外实验。
aGPCR的异常调节是和人类疾病高度相关的。海上靶标其中,最明显的是ADGRE2的突变会引起受体活化,肥大细胞的激活,导致振动性荨麻疹。ADGRE2在嗜中性粒细胞的促炎反应发挥作用,抑制它的功能会导致肝硬化。ADGRG1的突变或者在脑神经祖细胞的表达异常会导致BFPP,是一种以皮层发育不完全为主要特征的神经疾病。一些证据表明ADGRG6的变异和青少年脊柱侧弯相关。而就ADGRG1而言,通过对病人进行基因组测序,9个密码子关联的突变分布在整个蛋白上,这些突变会影响NTF和胶原蛋白III的结合,自催化水解以及细胞膜定位等生物学过程。因此,ADGRG1是所有aGPCR中和疾病关联研究最多的成员,也是最早选择作为治疗靶点的蛋白。
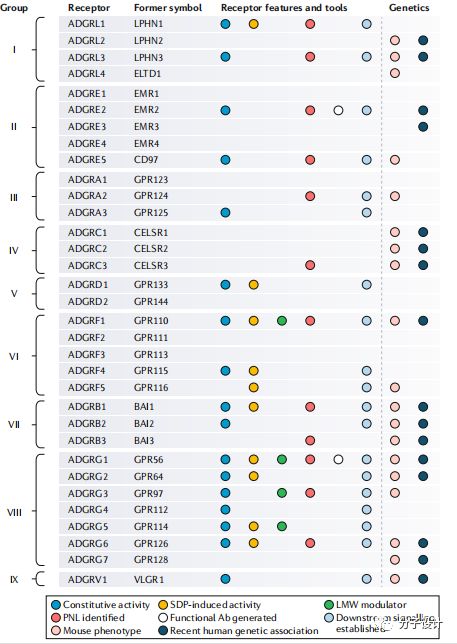
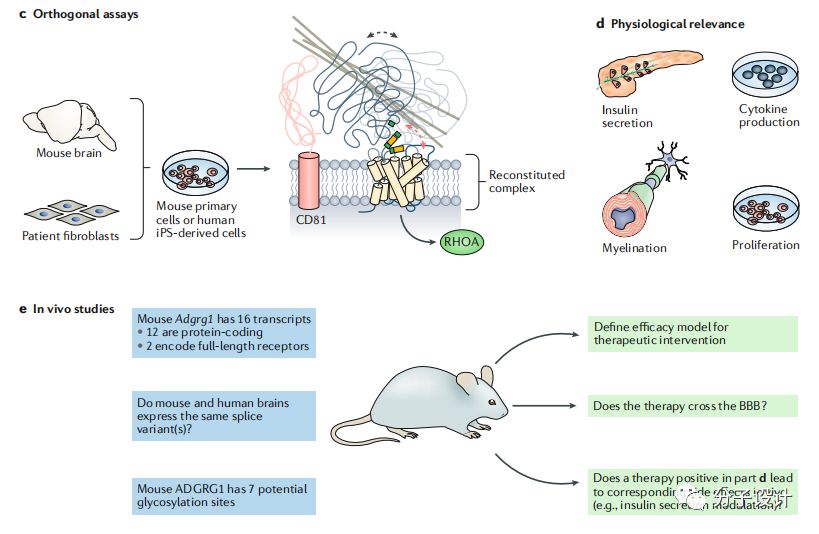
在小鼠上,aGPCR的基因改变也会产生一些和人类疾病相关的表型。但相比于人类,小鼠中和aGPCR相关联的基因更多,由此一些受体的改变会导致许多不同组织的表型改变。比如,ADGRB1的突变和小脑颗粒神经元前体的增殖,肌肉再生以及巨噬细胞吞噬相关。
在缺乏合适的小分子进行体内实验的情况下,将药物靶点在小鼠体内过表达是研究靶点活化对于人类健康影响的一种有效途径。实际上,在很多特定组织中aGPCR都是过表达的。ADGRB1在小鼠结肠中过表达可以阻止炎症发生和凋亡细胞的数量,而这两个表型在ADGRB1 KO小鼠中都明显增加,这些证据表明靶向改受体的激动剂可以治疗肠道炎症疾病。而ADGRG1在骨髓细胞中的过表达会引起急性髓性白血病,这时需要设计靶向的拮抗剂的方法了。虽然在小鼠体内小的框内缺失都会引起剧烈的表型改变,但是在体内探究aGPCR的方法还是需要进一步优化。比如,海上靶标把斑马鱼的ADGRA2的N端的亮氨酸富集区的重复片段去掉一个以后会导致鱼被根部神经发育障碍。比常规的基因破坏(敲除,敲入等)更精确的基因分析才能对aGPCR的作用方式提供更深入的理解。
体外细胞表型模型是比标准的信号转导检测更复杂也更难建立的实验方法但也更贴近哺乳动物的生理状态,在新药研发早期能提供有力的证据。而aGPCR相比于其他GPCR家族成员有独特的优势,因为aGPCR可以被Stachel的衍生肽(SDP)激动,因此可以基于此建立表型筛选体系发现有潜力的化合物。由于所有的aGPCR都会被SDP激动,因此也为新靶点的发现提供便利。考虑到Stachel的高度同源性,需要构建一个肽工具包才能激动所有成员,并通过改造原始肽来发现选择性激动剂或拮抗剂。即使产生阴性结果,这样的工具对于aGPCR的药物研发项目也是很有意义的。还有一点,如何确定该类家族以何种状态发挥靶向生理效应也是目前还没解决的问题,因为不是每个GPCR都是通过G蛋白发挥信号转导作用的。如果要探究G蛋白信号转导功能,目前是有很多工具的,比如毛喉素,百日咳毒素以及Gαq抑制剂。但是如果要探究其他作用机制,就要构建截短或突变的aGPCR来确认自催化裂解是否是蛋白必须的功能以及截去N端对于蛋白的生理功能的影响。其他方法还包括表达近跨膜区的N端部分或者重组N端。另外,aGPCR的有些功能需要蛋白的各个组分共同完成,而有一些则只需要某些部分。比如,只有ADGRG2的的C端才和钙感应蛋白相互作用并引起细胞内效应,而N端则不能。
明确受体激活的程度是至关重要的,因为这最终决定了发现小分子化合物的几率以及先导化合物或生物分子成为潜在药物的可能性。在有些例子中,GPCR上由人工导入或自发的突变会产生固有活性,这能有利于体系筛选。aGPCR同样如此,尤其将N端截去,aGPCR本身会有很高的活性。Stachel的存在对于研究下游信号转导是一个很好的工具,同时也能推动检测手段的发展。目前,对于aGPCR的信号转导机制的理解都是来源于Stachel的激动效应,也就是aGPCR的GPS发生自催化使得Stachel可以释放并暴露而被检测。比如,在ADGRE2中,提高N端和C端的解离能力可以使受体更容易被激活并促进肥大细胞的脱颗粒作用。然而在果蝇中,神经细胞的牵张敏感性并不需要受体的裂解,但基于Stachel的激活机制却会参与其中。对ADGRE2的信号传递机制的新认识孕育了一些广泛使用的GPCR检测手段,比如ERK的磷酸化,这些手段可以用于发现新的ADGRE2的调控分子。
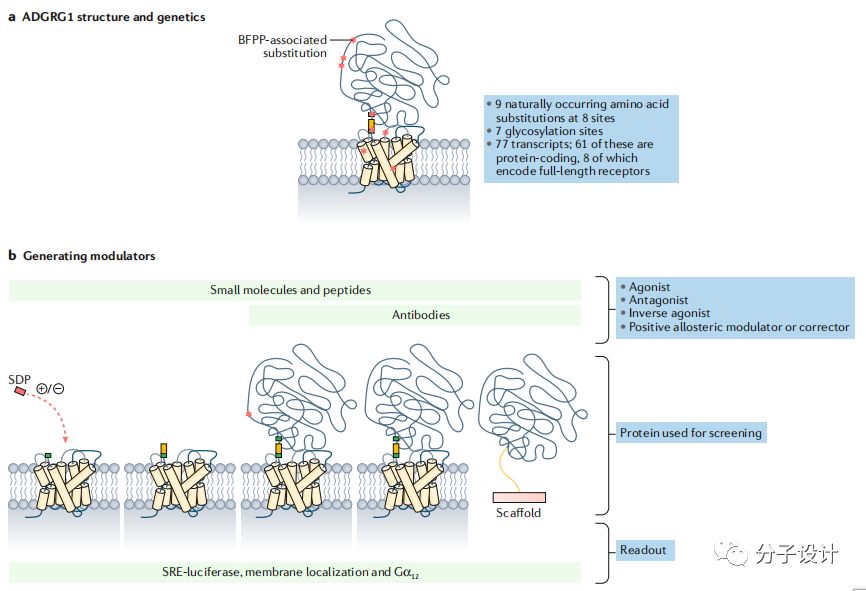
越来越多的研究人员也尝试用C端作为模型,因为重组的aGPCR很难表达。比如,Tall开发了一种利用C端和失活突变的Stachel来选择性激活蛋白的实验方法。Stachel的衍生肽(SDP)一般会形成β链并且是疏水的,这并不利于检测。然而,截短肽的长度也可以保留肽的活性。目前报道的SDP肽活性差别很大,所以留给肽的改造还有很大空间,只是目前还没有一个系统的SDP肽优化的方法。在使用基于Stachel的检测实验时需要注意的是有些aGPCR的激活是不依赖Stachel肽的。例如,ADGRG1可以直接被抗体类似物激活,不引起Stachel的信号传递。突触酰胺是大脑的代谢产物可以促进神经再生,它通过结合N端来激动ADGRF1,而自裂解的过程对于受体不是必须的。不依赖Stachel的ADGRG1的固有活性可以通过转化生长因子-β(TGF-β)脱落实验和血清反应元件(SRE)-荧光素酶实验检测。测定缺少Stachel介导的受体激活能力将会需要不同的检测平台。
aGPCR的N端不仅可以结合低分子量的小分子同样也可以结合抗体来改变的受体功能。抗体类似物和ADGRG1结合后可以产生不同的药力学性质包括拮抗和激动。这些抗体类似物发挥功能的机制是不依赖受体自裂解过程的,因为没有自裂解功能的受体同样也能被激活。目前有报道多种抗体能激活aGPCR,他们可能都是通过和N端结合来产生效应。这些抗体可以依照靶向鸦片类受体的方法来筛选拮抗剂和变构调控分子。这样的方法对于基于SDP的检测技术也是一个补充。抗体还可以通过空间位阻来阻断天然小分子的结合来破坏受体的信号转导。比如,在原代细胞检测实验中,对于ADGRGF1,抗N端抗体可以阻断由突触酰胺引起的cAMP信号。这样的抗体也可以用来测定结构和基于结构的药物设计。
最后,靶向N端的抗体在小鼠关节炎模型中被证明是有治疗潜力的,这样的抗体是需要ADGRE5介导的信号通路的参与,而这和其他基于抗体的机制是不同的。此外,靶向N端还可以选择性地调控aGPCRs下游不同的信号通路。例如,ADGRG2可以激活NF- κB和cAMP,但自裂解过程只能诱导cAMP信号。Gαq的抑制剂可以部分抑制ADGRG2的NF- κB信号,这表明可能非Gαq或者说是非G蛋白部分也参与介导了NF- κB的激活。
N端还可以被翻译后修饰,比如糖基化,有些受体可能含有25个糖基化位点。ADGRE5上的N端分布的糖基化作用对于配体CD55的结合是有利的。因此任何可以靶向ADGRE5的N端的检测技术都应该在被化学修饰的细胞环境下进行。在人类肿瘤来源的细胞系中,抗体表位包括糖基化位点可以差异性地识别ADGRE5,这也验证了使用该类细胞体系的重要性。
另一个靶向N端的方法就是抑制受体的自裂解过程,从而可以潜在抑制Stachel介导的激活机制。但是由于自裂解过程是发生在细胞内的,所以抑制剂分子需要穿过双层细胞膜来发挥活性。而抗自裂解的突变会产生完全不同的细胞反应,这也给这一类方法带来了不确定性。
在生理条件下,Stachel和受体的紧密结合很好地解决了脱靶的问题,但是在设计aGPCR的活化肽的时候,特异性一直是一个需要面对的问题。Stachel的序列在33个人源aGPCR中都是很保守的,并且实验证据表明一个SDP可以激活多个aGPCR成员。因此,在进行药物发现时,测试SDP在其他aGPCR受体的结合能力是很有必要的。另外,确认SDP是否以细胞穿模肽TAT相同的方式通过血脑屏障对于将该类肽用于治疗中枢神经系统疾病是很有意义的。
对于鸦片受体,由受体内化而产生的快速耐药反应是早期药物发现的一个问题,aGPCR同样如此。这类受体的内化机制通常是依赖β-arrestin的。然而,受体以什么样的速率进行循环是一个动力学的问题,也决定了快速耐药反应的程度。当把ADGRF5在小鼠体内KO之后,ADGRF5的基因缺陷表型出现时间很短,表明该基因有很快的循环过程。快速耐药反应可能会影响其他信号通路和机制而导致意想不到的结果。
近年来对aGPCR的进一步认识,尤其是在机制识别方面的突破,提示开发能调控aGPCR功能的活性分子对于该类受体的药物发现是具有里程碑意义的。然而,考虑到该类家族的特殊性,包括复杂的N端区域以及信号传导机制,我们预期在aGPCR药物发现领域会有更多的机遇与挑战。